
Kingdom Animalia Family Hominidae Rank Species Phylum Chordata | Suborder Haplorhini Genus †Australopithecus Higher classification Southern ape Order Primate | |
 | ||
Scientific name Australopithecus afarensis Similar Southern ape, Great apes, Australopithecus africanus, Homo habilis, Paranthropus boisei | ||
Australopithecus afarensis (Latin: "Southern ape from Afar") is an extinct hominin that lived between 3.9 and 2.9 million years ago. A. afarensis was slenderly built, like the younger Australopithecus africanus. A. afarensis is thought to be more closely related to the genus Homo (which includes the modern human species Homo sapiens), whether as a direct ancestor or a close relative of an unknown ancestor, than any other known primate from the same time. Some researchers include A. afarensis in the genus Praeanthropus.
Contents
- Localities
- Craniodental features and brain size
- Skeletal morphology and locomotion
- Behaviour
- Specimens of A afarensis
- Related work
- References

The most famous fossil is the partial skeleton named Lucy (3.2 million years old) found by Donald Johanson and colleagues, who, in celebration of their find, repeatedly played the Beatles song Lucy in the Sky with Diamonds.

Localities
Australopithecus afarensis fossils have only been discovered within Eastern Africa. Despite Laetoli being the type locality for A. afarensis, the most extensive remains assigned to the species are found in Hadar, Afar Region of Ethiopia, including the above-mentioned "Lucy" partial skeleton and the "First Family" found at the AL 333 locality. Other localities bearing A. afarensis remains include Omo, Maka, Fejej, and Belohdelie in Ethiopia, and Koobi Fora and Lothagam in Kenya.
Craniodental features and brain size

Compared to the modern and extinct great apes, A. afarensis has reduced canines and molars, although they are still relatively larger than in modern humans. A. afarensis also has a relatively small brain size (about 380–430 cm3) and a prognathic face (i.e. a face with forward-projecting jaws).
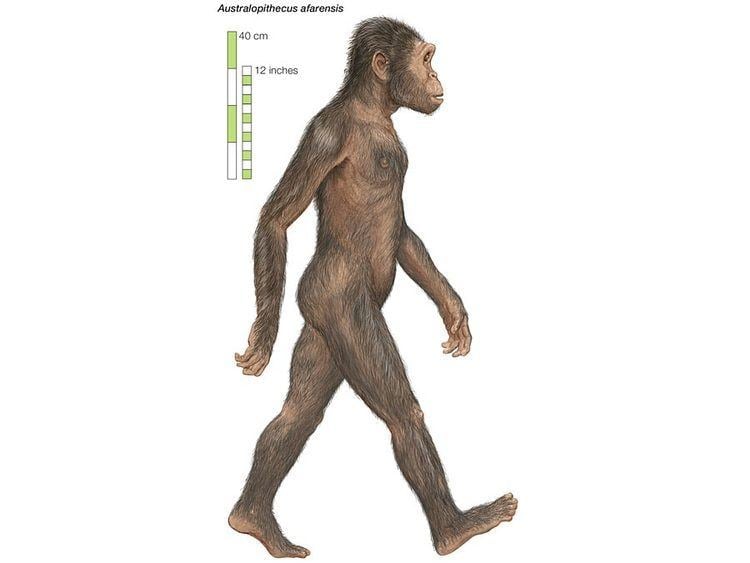
Skeletal morphology and locomotion

Considerable debate surrounds the locomotor behaviour of A. afarensis. Some studies suggest that A. afarensis was almost exclusively bipedal, while others propose that the creatures were partly arboreal. The anatomy of the hands, feet, and shoulder joints in many ways favour the latter interpretation. In particular, the morphology of the scapula appears to be ape-like and very different from modern humans. The curvature of the finger and toe bones (phalanges) approaches that of modern-day apes, and is suggestive of their ability to efficiently grasp branches and climb. Alternatively, the loss of an abductable great toe and therefore the ability to grasp with the foot (a feature of all other primates) suggests A. afarensis was no longer adapted to climbing.
A number of traits in the A. afarensis skeleton strongly reflect bipedalism, to the extent some researchers have suggested bipedality evolved long before A. afarensis. In overall anatomy, the pelvis is far more human-like than ape-like. The iliac blades are short and wide, the sacrum is wide and positioned directly behind the hip joint, and evidence of a strong attachment for the knee extensors is clear. While the pelvis is not wholly human-like (being markedly wide, or flared, with laterally oriented iliac blades), these features point to a structure that can be considered radically remodeled to accommodate a significant degree of bipedalism in the animals' locomotor repertoire.
Importantly, the femur also angles in toward the knee from the hip. This trait would have allowed the foot to have fallen closer to the midline of the body, and is a strong indication of habitual bipedal locomotion. The feet also feature adducted big toes, making it difficult if not impossible to grasp branches with the hindlimbs. The loss of a grasping hindlimb also increases the risk of an infant being dropped or falling, as primates typically hold onto their mothers while the mother goes about her daily business. Without the second set of grasping limbs, the infant cannot maintain as strong a grip, and likely had to be held with help from the mother. The problem of holding the infant would be multiplied if the mother also had to climb trees. Bones of the foot (such as the calcaneus) also indicate bipedality.
Computer simulations using dynamic modeling of the skeleton's inertial properties and kinematics suggest A. afarensis was able to walk in the same way modern humans walk, with a normal erect gait or with bent hips and knees, but could not walk in the same way as chimpanzees. The upright gait would have been much more efficient than the bent knee and hip walking, which would have taken twice as much energy.
A. afarensis probably was quite an efficient bipedal walker over short distances, and the spacing of the footprints at Laetoli indicates they were walking at 1.0 m/s or above, which matches human small-town walking speeds. Yet, this can be questioned, as finds of Australopithecus foot bones indicate the Laetoli footprints may not have been made by Australopithecus. Many scientists also doubt the suggestion of bipedalism, and argue that even if Australopithecus really did walk on two legs, it did not walk in the same way as humans.
The presence of a wrist-locking mechanism, though, might suggest they engaged in knuckle-walking. (However, these conclusions have been questioned on the basis of close analysis of knuckle-walking and the comparison of wrist bones in different species of primates). The shoulder joint is also oriented much more cranially (i.e. towards the skull) than that in modern humans, but similar to that in the present-day apes. Combined with the relatively long arms A. afarensis is thought to have had, this is thought by many to be reflective of a heightened ability to use the arm above the head in climbing behaviour. Furthermore, scans of the skulls reveal a canal and bony labyrinth morphology, which is not supportive to proper bipedal locomotion.
Upright bipedal walking is commonly thought to have evolved from knuckle-walking with bent legs, in the manner used by chimpanzees and gorillas to move around on the ground, but fossils such as Orrorin tugenensis indicate bipedalism around 5 to 8 million years ago, in the same general period when genetic studies suggest the lineage of chimpanzees and humans diverged. Modern apes and their fossil ancestors show skeletal adaptations to an upright posture used in tree-climbing, and upright, straight-legged walking has been proposed to have originally evolved as an adaptation to tree-dwelling.
Studies of modern orangutans in Sumatra have shown these apes use four legs when walking on large, stable branches and when swinging underneath slightly smaller branches, but are bipedal and maintain their legs very straight when using multiple small, flexible branches under 4 cm in diameter, while also using their arms for balance and additional support. This enables them to get nearer to the edge of the tree canopy to grasp fruit or cross to another tree.
Climate changes around 11 to 10 million years ago affected forests in East and Central Africa, establishing periods where openings in forest lands prevented travel through the tree canopy. During such times proto-hominins could have adopted upright walking behaviour for ever-increasing ground travel, while the ancestors of gorillas and chimpanzees continued to specialize in climbing vertical tree trunks and lianas with a bent-hip and bent-knee posture—which ultimately lead them also to adopt knuckle-walking for minimal ground travel. This differential development within the larger hominid community would result in A. afarensis being adapted to upright bipedalism for extensive ground travel while still using arms well adapted for climbing smaller trees. Still, the proto-hominins and the ancestors of chimpanzees and gorillas were the closest of relatives, and they shared anatomical features including a fused wrist bone, which may suggest that knuckle-walking was used for a time by human ancestors.
Other studies suggest an upright spine and a primarily vertical body plan in primates dates back to Morotopithecus bishopi in the Early Miocene of 21.6 Mya, the earliest human-like primates. Known from fossil remains found in Africa, australopithecines represent the group from which the ancestors of modern humans emerged. As generally used, the term "australopithecine" covers all early hominin fossils dated from about 7 million to 2.5 million years ago—and some others dated later, i.e., from 2.5 million to 1.4 million years ago. Australopithecines became extinct after that time.
Behaviour
Reconstruction of the social behaviour of extinct fossil species is difficult, but their social structure is likely to be comparable to that of modern apes, given the average difference in body size between males and females (sexual dimorphism). The degree of sexual dimorphism between males and females of A. afarensis is considerably debated. Some propose that males were substantially larger than females. If observations on the relationship between sexual dimorphism and social group structure from modern great apes are applied to A. afarensis, then these creatures most likely lived in small family groups containing a single dominant male and a number of breeding females. Other studies have shown there could have been substantial overlap between males and females in size. This, along with the reduction of the canine teeth, has been argued to suggest A. afarensis males and females were monogamous. Males may have engaged in provisioning behavior, and the need for carrying may have led to the evolution of bipedalism.
For a long time, no known stone tools were associated with A. afarensis, and paleoanthropologists commonly thought stone artifacts only dated back to about 2.5 Mya. However, a 2010 study suggests the hominin species ate meat by carving animal carcasses with stone implements. This finding pushes back the earliest known use of stone tools among hominins to about 3.4 Mya.
Specimens of A. afarensis
Related work
Further findings at Afar, including the many hominin bones in site 333, produced more bones of concurrent date, and led to Johanson and White's eventual argument that the Koobi Fora hominins were concurrent with the Afar hominins. In other words, Lucy was not unique in evolving bipedalism and a flat face.
In 2001, Meave Leakey proposed a new genus and species, Kenyanthropus platyops, for fossil cranium KNM WT 40000. The fossil cranium appears to have a flat face, but the remains are heavily fragmented. It has many other characteristics similar to A. afarensis remains. It is so far the only representative of its species and genus, and the timeframe for its existence falls within that of A. afarensis.
Another species, called Ardipithecus ramidus, was found by Tim White and colleagues in 1992. This was fully bipedal at 4.4 to 5.8 Mya, yet appears to have been contemporaneous with a woodland environment. Scientists have a rough estimation of the cranial capacity of Ar. ramidus at 300 to 350 cm³, from the data released on the Ardipithecus specimen nicknamed Ardi in 2009.