
 | ||
Antigen presentation describes a vital immune process. Because T cells only recognise antigens displayed on the cell surfaces, an antigen-presenting complex is needed for detecting infectious cells. After infection with viruses or bacteria, the cells present endogenous peptide fragments derived from the pathogen by major histocompatibility complex (MHC) molecules on the cell surface. There are two types of MHC, depending on the source of the antigens: MHC class I molecules (MHC-I) binds peptides from the cytosol, while peptides generated in vesicles are bound to MHC class II (MHC-II). T cells can recognise among ten to hundreds of thousands peptides because each MHC molecule can bind a different range of peptides.
Contents
Intracellular antigens: Class I
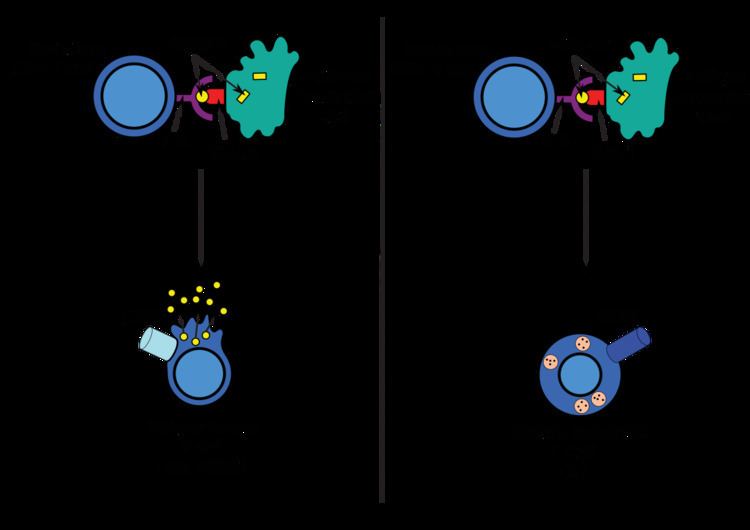
Cytotoxic T cells regularly patrol body cells to maintain the organism healthy. Whenever they encounter signs of disease, caused for example by the presence of viruses or intracellular bacteria, they initiate processes to destroy the potentially harmful cell. To effect recognition, all nucleated cells in the body (along with platelets) display class I major histocompatibility complex (MHC-I molecules). Antigens generated endogenously within the cells (DAMPs) are bounded to MHC-I molecules and presented on the cell surface. This antigen presentation pathway enables the immune system to detect transformed or infected cells displaying peptides from modified-self or foreign proteins.
In the presentation process, these proteins are mainly degraded into small peptides by cytosolic proteases in the proteasome, but there are also other cytoplasmic proteolytic pathways. Then, the transporter associated with antigen processing (TAP) translocates the cytosolic peptides into the ER lumen in an ATP-dependent transport mechanism. There are several ER chaperones involved in MHC-I assembly, as calnexin, clareticulin and tapasin. Because of the association between TAP and MHC-I, the peptides are loaded to MHC-I molecules. After releasing from tapasin, peptide-MHC-I complexes (pMHC-I) exit the ER and are transported to the cell surface by exocytic vesicles.
Naïve anti-viral CD8+ T cells (TCD8+) cannot directly eliminate transformed or infected cells. They have to be activated by the pMHC-I complexes of antigen-presenting cells (APCs) . Antigen can be presented directly or indirectly (cross presentation) from virus-infected and non-infected cells. After the interaction between pMHC-I and TLR, in presence of costimulatory signals and/or cytokines,T cells are activated and kill the target cells (infected or damaged cells) by inducing cytotoxicity.
Cross-presentation is a special case in which MHC-I molecules are able to present extracellular antigens, usually displayed by MHC-II molecules. This ability appears in several APCs, mainly dendritic cells, that stimulate CD8+ T cells. This process is essential when APCs are not directly infected, triggering antiviral and antitumour immune responses.
Extracellular antigens: Class II
Antigens from the extracellular space (PAMPs) and also endogenous ones, are enclosed into vesicles and presented on the cell surface by MHC-II molecules to CD4+ T cells. Only professional APCs have this class of MHC molecules, such as dendritic cells, B cells or macrophages, so expression of MHC-II molecules is more restricted than MHC-I.
APCs usually internalize exogenous antigens by endocytosis, but also by pinocytosis, macroautophagy, endosomal microautophagy or chaperone-mediated autophagy. In the first case, after internalisation, the antigens are enclosed in vesicles called endosomes. There are three compartements involved in this antigen presentation pathway: early endosomes, late endosomes or endolysosomes and lysosomes, where antigens are hydrolized by lysosome-associated enzymes (acid-dependent hydrolases, glycosidases, proteases, lipases...). This process is favored by the gradually reduction of the pH. The main protease is cathepsin and the result is the degradation of the antigens into oligopeptides.
MHC-II molecules are transported from the ER to the endosomes by the protein Invariant chain (Ii, CD74). A non classical MHC-II molecule (HLA-DM) catalyses the exchange of part of the CD74 (CLIP) with the peptide antigen. Peptide-MHC-II complexes (pMHC-II) are transported to the plasma membrane and the processed antigen is presented to CD4+ T cells in the lymph nodes.
DCs undergo a process of maturation while migrating, via chemotactic signals, to lymphoid tissues, in which they lose the phagocytic capacity and develop an increased ability to communicate with T-cells by antigen-presentation. As well as in CD8+ T cells, APCs need pMHC-II and aditional costimulatory signals to activate T helper cells.
Intact antigens: Presentation to B cells
B-cell receptors bind to intact, undigested antigens, rather than to proteins which have been digested and presented by MHC molecules. Small antigens sometimes encounter B cells after diffusing on their own to lymph nodes, but larger pieces of intact antigen appear to be presented to B cells by follicular dendritic cells and macrophages. It seems that these macrophages express comparatively lower levels of lysosomal enzymes and thus are less likely to digest the antigen they have captured before presenting it.