
 | ||
Cellular senescence is the phenomenon by which normal diploid cells cease to divide. In culture, fibroblasts can reach a maximum of 50 cell divisions before becoming senescent. This phenomenon is known as "replicative senescence", or the Hayflick limit. Replicative senescence is the result of telomere shortening that ultimately triggers a DNA damage response. Cells can also be induced to senesce via DNA damage in response to elevated reactive oxygen species (ROS), activation of oncogenes and cell-cell fusion, independent of telomere length. As such, cellular senescence represents a change in "cell state" rather than a cell becoming "aged" as the name confusingly suggests. Nonetheless, the number of senescent cells in tissues rises substantially during normal aging.
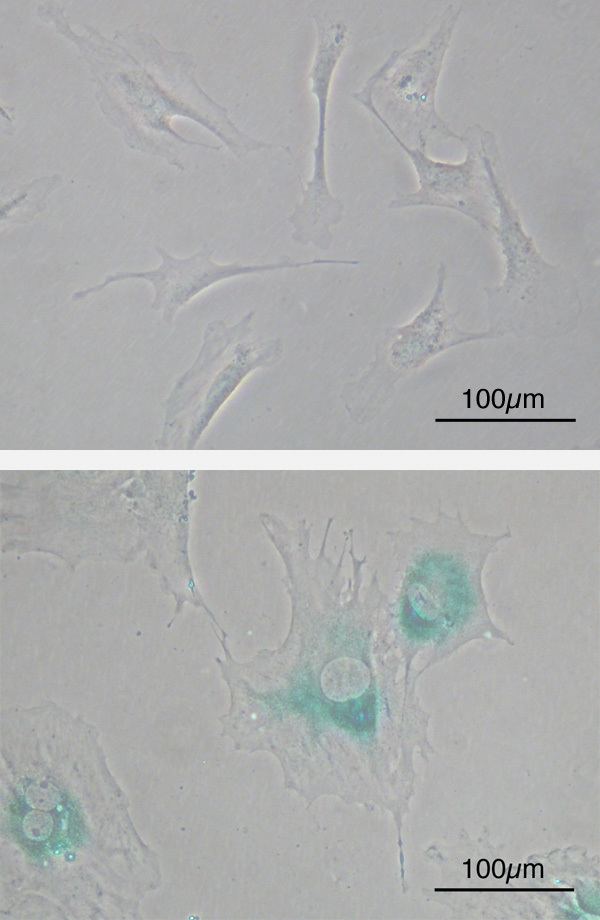
Although senescent cells can no longer replicate, they remain metabolically active and commonly adopt an immunogenic phenotype consisting of a pro-inflammatory secretome, the up-regulation of immune ligands, a pro-survival response, promiscuous gene expression (pGE) and stain positive for senescence-associated β-galactosidase activity. Senescence-associated beta-galactosidase, along with p16Ink4A, is regarded to be a biomarker of cellular senescence. Nonetheless, false positives exist for maturing tissue macrophages and senescence-associated beta-galactosidase as well as for T-cells p16Ink4A.
A Senescence Associated Secretory Phenotype (SASP) consisting of inflammatory cytokines, growth factors, and proteases is another highly characteristic feature of senescent cells. SASP contributes to many age-related diseases, including type 2 diabetes and atherosclerosis. The damaging effects of SASP have motivated researchers to develop senolytic chemicals that would kill and eliminate senescent cells to improve health in the elderly. Healthy mice treated with senolytics have shown improved cardiac and vascular, function. Removal of senescent cells in normal mice increased healthspan as well as life expectancy,
The nucleus of senescent cells is characterized by senescence-associated heterochromatin foci (SAHF) and DNA segments with chromatin alterations reinforcing senescence (DNA-SCARS). Senescent cells affect tumour suppression, wound healing and possibly embryonic/placental development and a pathological role in age-related diseases.
The experimental elimination of senescent cells from transgenic progeroid mice and non-progeroid, naturally-aged mice led to greater resistance against aging-associated diseases.
Moreover, cellular senescence is not observed in several organisms, including perennial plants, sponges, corals, and lobsters. In those species where cellular senescence is observed, cells eventually become post-mitotic when they can no longer replicate themselves through the process of cellular mitosis; i.e., cells experience replicative senescence. How and why some cells become post-mitotic in some species has been the subject of much research and speculation, but (as noted above) it is sometimes suggested that cellular senescence evolved as a way to prevent the onset and spread of cancer. Somatic cells that have divided many times will have accumulated DNA mutations and would therefore be in danger of becoming cancerous if cell division continued. As such, it is becoming apparent that senescent cells undergo conversion to an immunogenic phenotype that enables them to be eliminated by the immune system.